
International Journal of Earth Science and Geophysics
(ISSN: 2631-5033)
Volume 5, Issue 1
Research Article
DOI: 10.35840/2631-5033/1824

Article Formats
Timing of Anomalously Rapid Ammonite Diversification and Extinction from the Upper Cretaceous (SantonianCampanian) of James Ross Island, Antarctica using the First Antarctic Magnetostratigraphy for this Time Interval
Table of Content
Figures

Figure 1: Geographic position of the Marambio...
Geographic position of the Marambio Group (Upper Cretaceous) sedimentary basin on the Antarctica Peninsula, and the positions of the measured sections used in this paper.
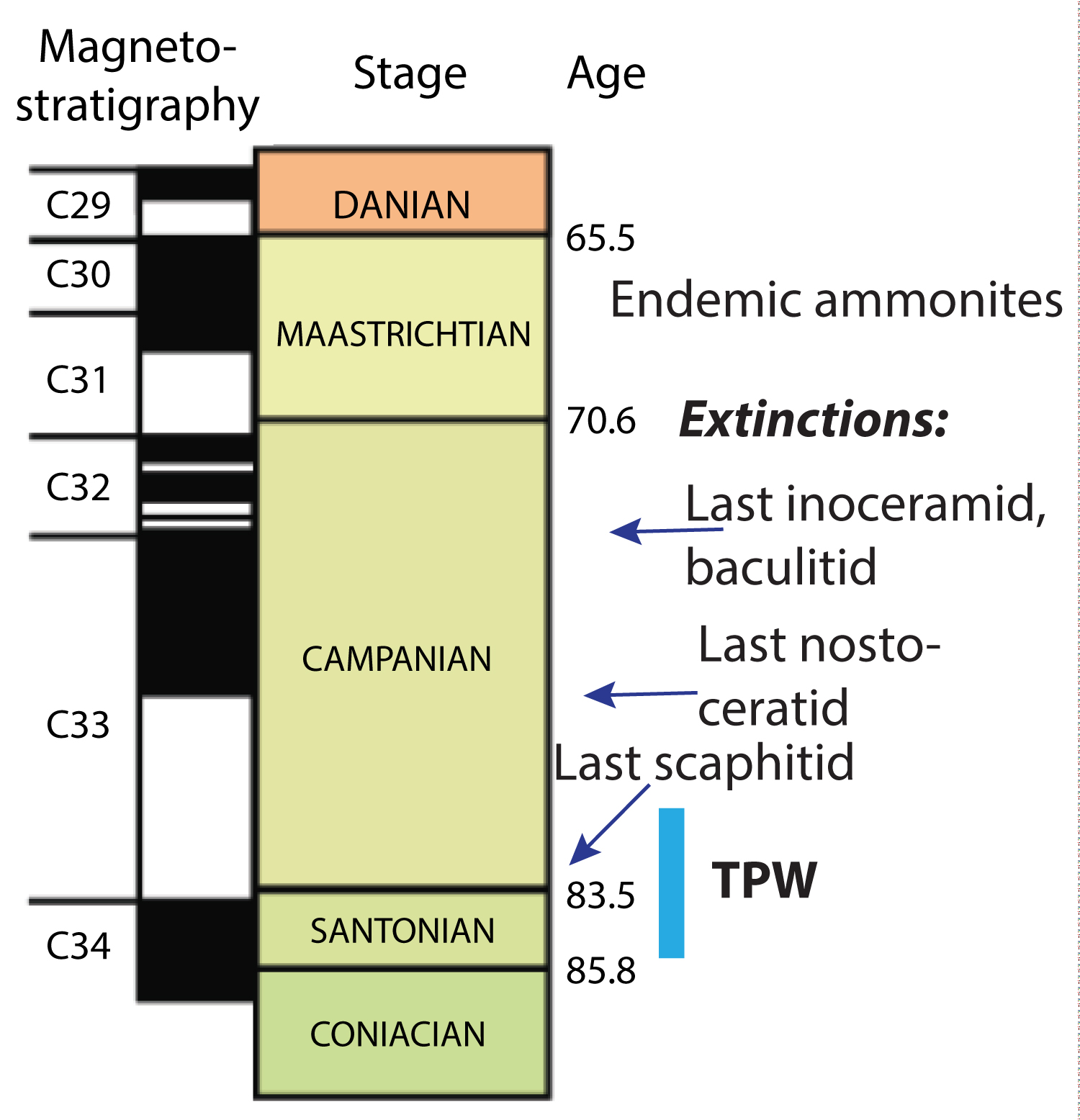
Figure 2: Times of early extinctions, James Ross...
Times of early extinctions, James Ross Island Basin. The period of True Polar Wander that has been previously recognized elsewhere on the globe during the Santonian into Early Campanian shown as blue bar to right of graph, marked TPW.

Figure 3: James Ross Island sections sampled for...
James Ross Island sections sampled for this paper, with lithostratigraphy, magnetostratigraphy and ranges of important ammonites. Each of the sections sampled is shown relative to its stratigraphic height as measured from low in the Hidden Lake Formation. The numbers to the right of each section refer to a single paleomagnetic sampling site (one or more cores taken and analyzed. The results from the core analyses are listed in Table 1. Biostratigraphic zones from prior work are listed to left of column (Zones I-VIII) of [6-8].
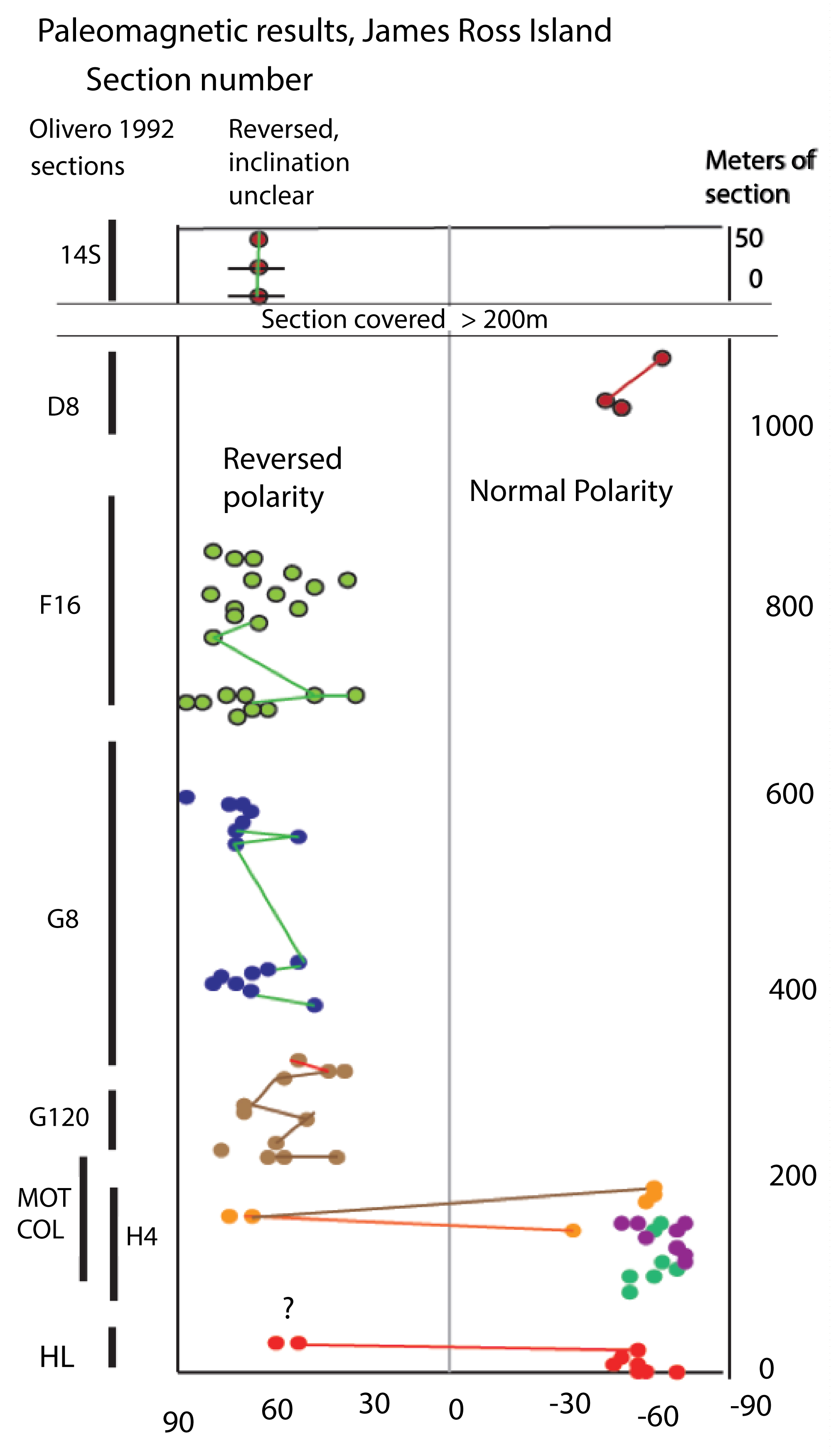
Figure 4: Primary paleomagnetic results from the...
Primary paleomagnetic results from the measured sections shown in Figure 3. The sections sampled shown on left. A long missing (covered) interval (not previously quantified) separates the top of section D8 and the base of 14S, as is also show in our Figure 1. This interval is at least 200m thick stratigraphically, and we interpret it to include much of the time of Chron 33N, as well as being correlative in age to Rabot Point, James Ross Island [15].
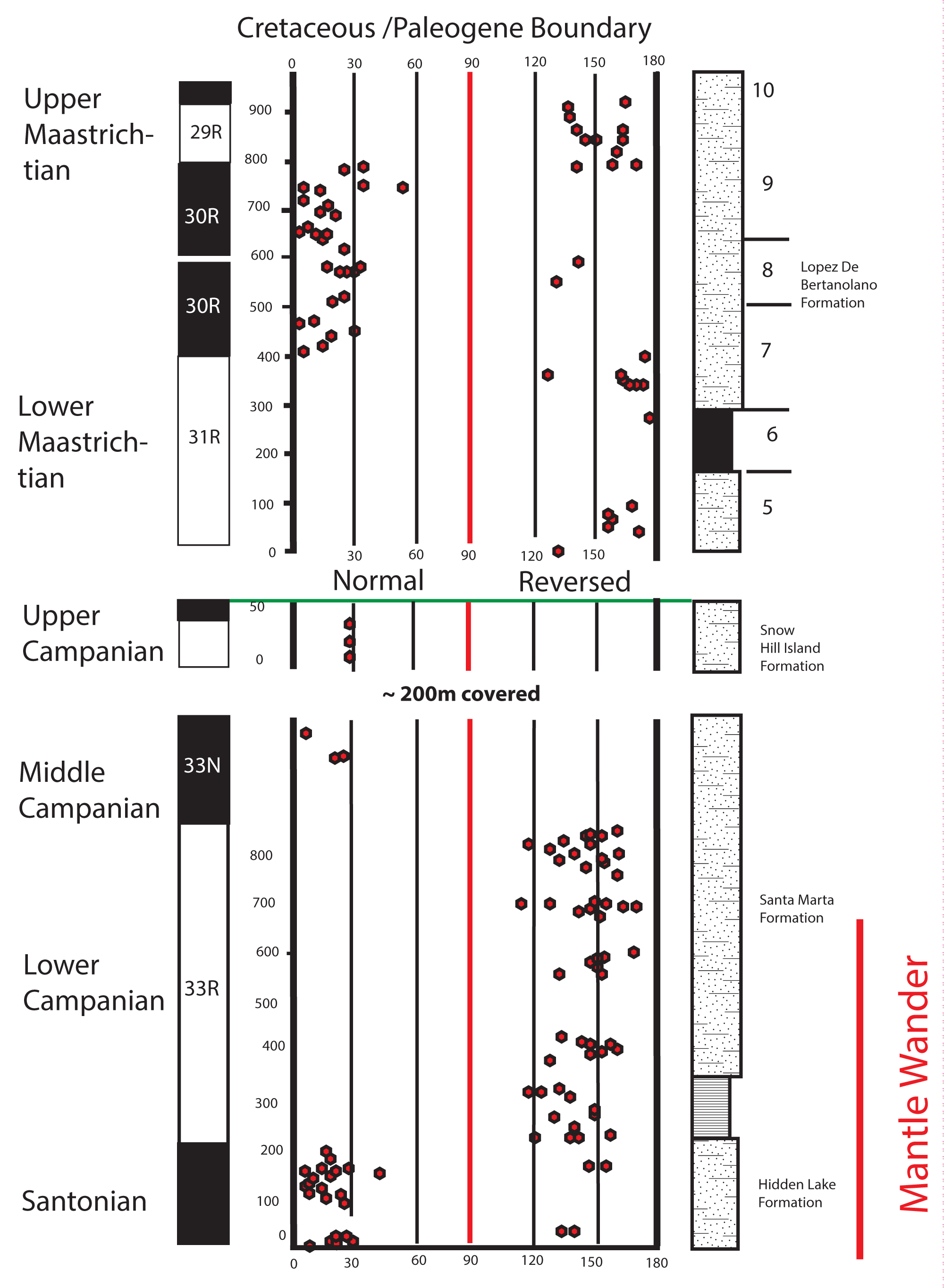
Figure 5: Combined magnetostratigraphy for the entire basin...
Combined magnetostratigraphy for the entire basin from Santonian through Maastrichtian, by combining the new results shown here with those previously shown in Tobin, et al. [1]. Numbers to right of measured section on upper half of graph correspond the "Molluscan Units" [4] of previous workers. The time interval of the possible True Polar Wander (labled here as "Mantle Wander", the second name given to the process) that has been elsewhere identified during the Santonian time interval shown in red to right bottom of graph.
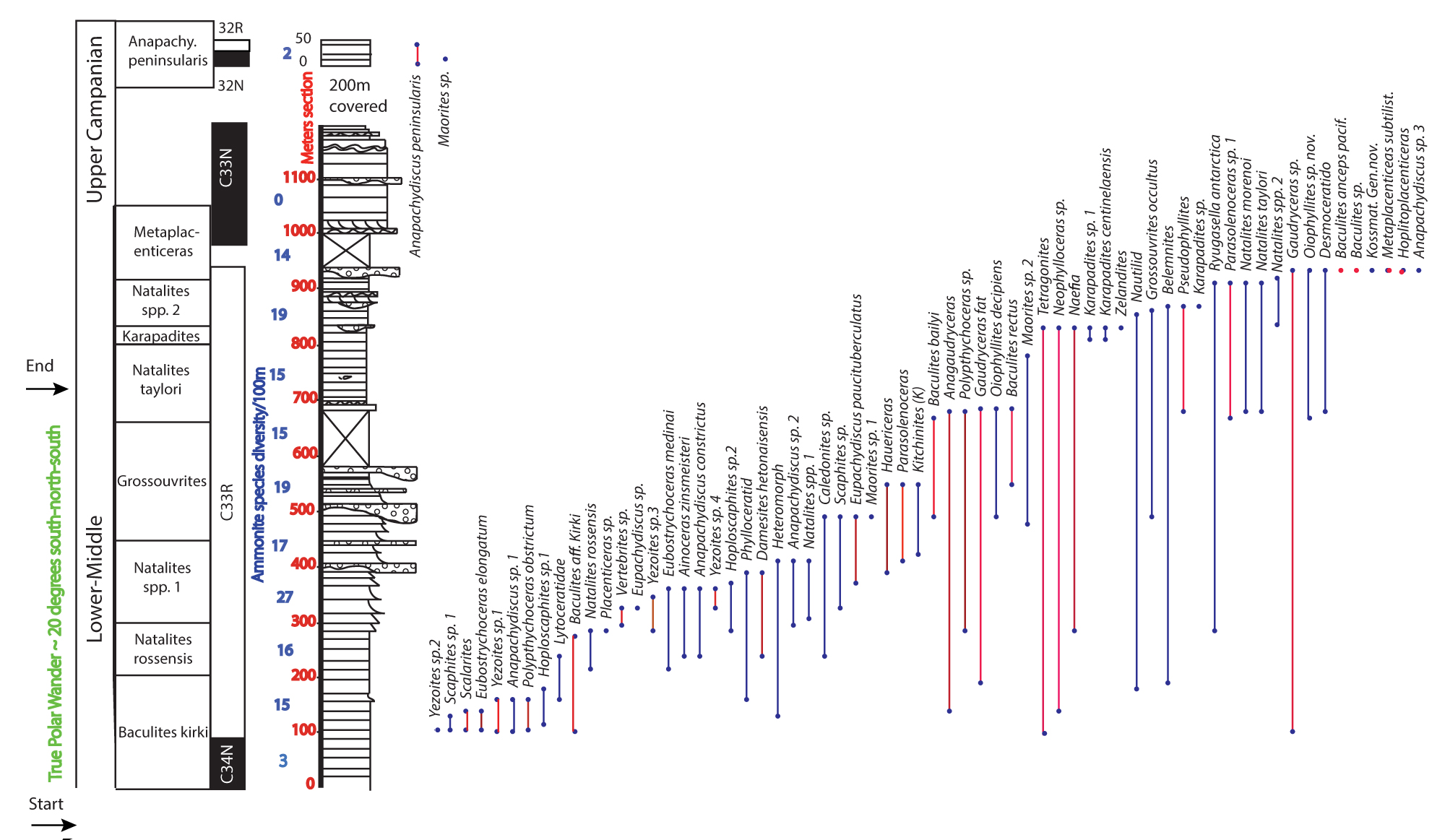
Figure 6: Ranges of ammonites from James Ross Island...
Ranges of ammonites from James Ross Island plotteed against sections. Ammonte species shown in red are shared with Indopacific Province ammonite species, [6-8]. Ranges of Indopacific ammonites come from [16]. The interval of True Polar Wander identified on far left of graph.
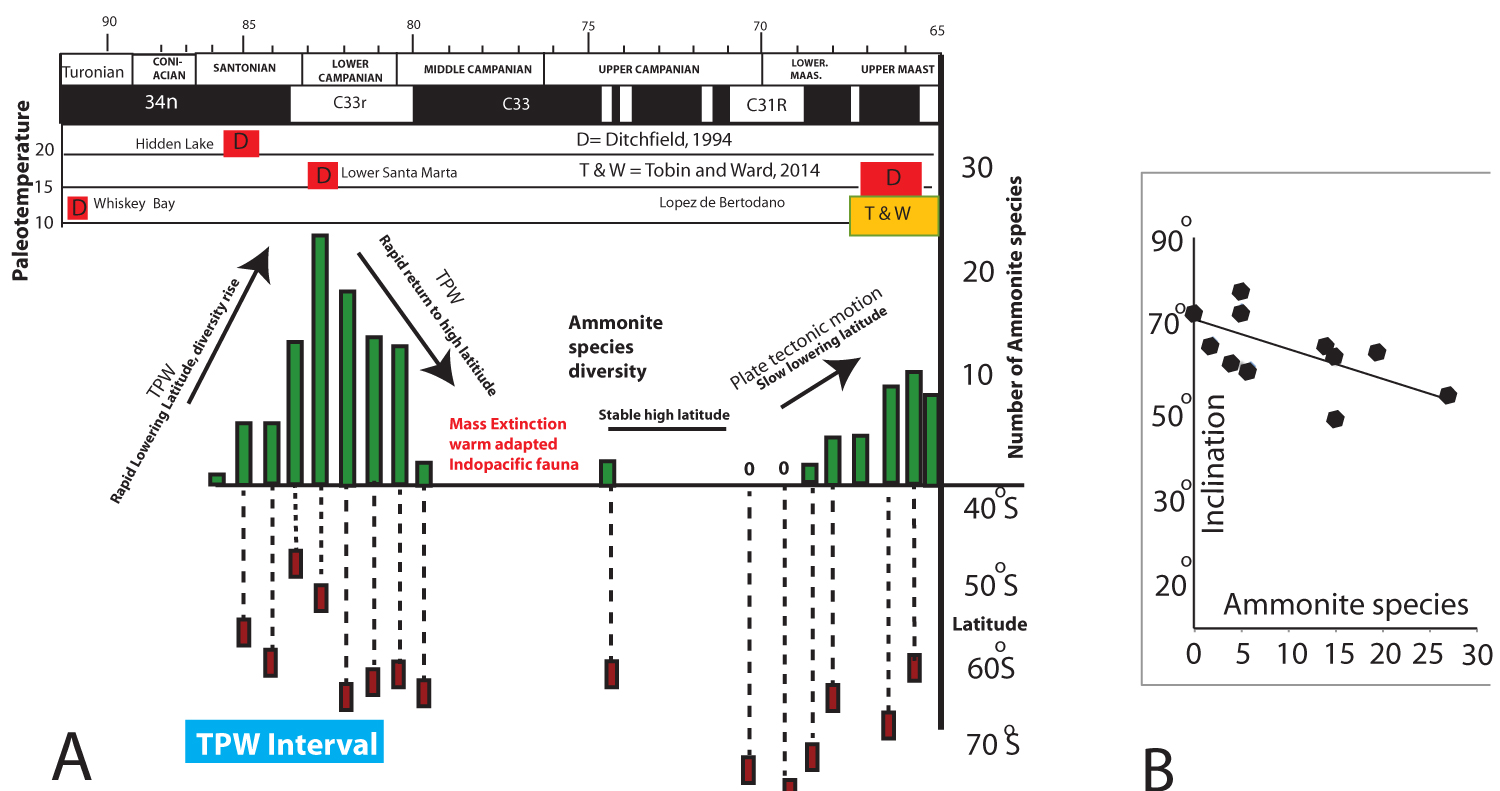
Figure 7: Summary figure integrating time, ammonite...
Summary figure integrating time, ammonite diversity, paleotemperature, and interpolated paleolatitude (from inclination data presented here and in Tobin, et al. [1] for the entire basin for the Late Cretaceous. There is a significant correlation between ammonite diversity and inclination, in line with ecological results from modern day organisms of higher diversity correlated to lower latitudes. Temperatures come from sources listed in the figure.

Figure 8: Graphic showing proposed TPW movement of...
Graphic showing proposed TPW movement of Antarctica, illustraing the relative geographic movement of Antarctica during the hypothesized True Polar Wander event in the Santonian.
References
- Tobin TS, Ward PD, Steig EJ, Olivero EB, Hilburn IA, et al. (2012) Extinction patterns, δ18 O trends, and magnetostratigraphy from a southern high-latitude Cretaceous-Paleogene section: Links with Deccan volcanism. Paleogeography Paleoclimatology Paleoecology 350-352: 180-188.
- Steinberger B, Torsvik TH (2008) Absolute plate motions and true polar wander in the absence of hotspot tracks. Nature 452: 620-623.
- Askin RA (1988) Campanian to paleocene palynological succession of seymour and adjacent islands, northeastern Antarctic Peninsula. In: Feldman RM, Woodburne MO, Geology and paleontology of seymour Island, Antarctic Peninsula. Geological Society of America, Memoir, 131-153.
- Macellari C (1986) Late campanian-maastrichtian ammonite fauna from Seymour Island (Antarctic Peninsula). Journal of Paleontology 60: 1-55.
- Elliot David H, Askin Rosemary A, Kyte Frank T, Zinsmeister William J (1994) Iridium and dinocysts at the cretaceous-tertiary boundary on seymour Island, Antarctica: Implications for the K-T event. Geology 22: 675-678.
- Olivero EB (1992) Asociaciones de Amonites de la Formacion Santa Marta (Cretacico Tardio), Isla James Ross. Geologia De La Isla James Ross 47-76.
- Olivero EB (2012) Sedimentary cycles, ammonite diversity and palaeoenvironmental changes in the Upper Cretaceous Marambio Group, Antarctica. Cretaceous Research 34: 348-366.
- Olivero EB, Medina FA (2000) Patterns of Late Cretaceous ammonite biogeography in southern high latitudes: The family Kossmaticeratidae in Antarctica. Cretaceous Research 21: 269-279.
- Olivero EB, Ponce JJ, Marsicano CA, Martinioni DR (2007) Depositional settings of the basal López de Bertodano Formation, Maastrichtian, Antarctica. Revista de laAsociación Geológica Argentina 62: 521-529.
- Crame JA (1982) Late Mesozoic bivalve biostratigraphy of the Antarctic Peninsula region. Journal of the Geological Society 139: 771-783.
- Crame JA, Francis JE, Cantrill DJ, Pirrie D (2004) Maastrichtian stratigraphy of Antarctica. Cretaceous Research 25: 411-423.
- Kirschvink JL (1980) The least-squares line and plane and the analysis of palaeomagnetic data. Geophys J R astr Soc 62: 699-718.
- Kirschvink Joseph L, Kopp Robert E, Raub Timothy D, Baumgartner Christopher T, Holt John W (2008) Rapid, precise, and high-sensitivity acquisition of paleomagnetic and rock-magnetic data: Development of a low-noise automatic sample changing system for superconducting rock magnetometers. Geochemistry Geophysics Geosystems 9.
- Crame JA, Pirrie D, Riding JB, Thomson MRA (1991) Campanian-Maastrichtian (Cretaceous) stratigraphy of the James Ross Island area, Antarctica. Journal of the Geological Society 148: 1125-1140.
- Milanese F, Olivero E, Kirschvink J, Rapalini A (2017) Magnetostratigraphy of the rabot formation, upper cretaceous, James ross basin, Antarctic peninsula. Cretaceous Research 72: 172-187.
- Ward P, Haggart J, Mitchell R, Kirschvink J, Tobin T (2012) Integration of macrofossil biostratigraphy and magnetostratigraphy for the Pacific Coast Upper Cretaceous (Campanian - Maastrichtian) of North America and implications for correlation with the Western Interior and Tethys. GSA Bulletin 124: 957-974.
- Ward P, Mitchell R, Haggart J, Catlin E (2015) Quantitative morphological description of the Late Cretaceous ammonite Baculites inornatus Meek from western North America: Implications for species concepts in the biostratigraphically important Baculitidae. Journal of Paleontology 89: 594-610.
- Mound J, Mitroveca J, G Milne (2000) Sea level change and true polar wander during the late cretaceous. Geophysical Research Letters 28: 2057-2060.
- Ditchfield PW, Marshall JD, Pirrie F (1994) High latitude palaeotemperature variation: New data from the Thithonian to Eocene of James Ross Island, Antarctica. Palaeogeography, Palaeoclimatology, Palaeoecology 107: 79-101.
- Barker PF, Thomas E (2004) Origin, signature and palaeoclimatic influence of the Antarctic Circumpolar Current. Earth Sci Rev 66: 143-162.
- Mitchell R, Ward P, J Haggart J. In Review. The end Cretaceous superchron identified in the Upper Cretaceous Great Valley Sequence of California. Cretaceous Research.
- Witts JD, Bowman VC, Wignall PB, Crame C, Francis JE, et al. (2015) Evolution and extinction of Maastrichtian (Late Cretaceous) cephalopods from the López de Bertodano Formation, Seymour Island, Antarctica. Palaeogeography, Palaeoclimatology, Palaeoecology 418: 193-212.
Author Details
Peter D Ward1*, Ross Mitchell2 and Eric J Steig3
1Department of Biology, University of Washington, USA
2Department Geology and Geophysics, Yale University, USA
3Department of Earth and Space Sciences, University of Washington, USA
Corresponding author
Peter D Ward, Department of Biology, University of Washington, Seattle, 98195 USA, Tel: 206-543-2962.
Accepted: April 18, 2019 | Published Online: April 20, 2019
Citation: Ward PD, Mitchell R, Steig EJ (2019) Timing of Anomalously Rapid Ammonite Diversification and Extinction from the Upper Cretaceous (Santonian-Campanian) of James Ross Island, Antarctica using the First Antarctic Magnetostratigraphy for this Time Interval. Int J Earth Sci Geophys 5:024.
Copyright: © 2019 Ward PD, et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Abstract
Searching for the causes and consequences of the Cretaceous-Paleogene mass extinction instigated decades of paleontological research on Seymour Island, Antarctica. However, biological changes, including rapid diversification followed by equally rapid extinctions of mollusks occurred more than 20 million years prior to the K/Pg boundary event in Antarctica, yet understanding these events has been hampered by the paucity of high-resolution correlation of what has been biostratigraphically identified as Santonian - through-Campanian Stage strata of the James Ross Basin (JRB) to the international stage stratotypes. This has been caused by endemism of ammonite species chosen as zonal indices, a lack of bentonite/ash age dating, and until recently, the absence of magnetostratigraphy. The publication by Tobin, et al. [1] produced a magnetostratigrpahic record for the Maastrichtian Stage on Seymour Island, but earlier strata, and other islands in the (also referred to as the James Ross Marambia Basin or MB) were not included. Here we complete the magnetostratigraphic record from the base of the Maastrichtian down to the Coniacian Stage.
This stratal (and time) interval coincides with a still unexplained, rapid increase in ammonite diversity beginning during the Santonian Age, and an equally rapid extinction of all of the newly appearing species by the mid-to late Campanian Age, as recognitized by chron boundaries presented here. The rapidity of both the Santonian Age diversification, and its mirror image reduction in diversity during the Campanian Age, is not explained through current understanding of Antarctic tectonics, paleoecology, and/or paleobiogeography. An alternative hypothesis can be found in invoking ecological effects caused by a previously reported episode of True Polar Wander, (or alternatively, Mantle Wander) a solid-body rotation of the planet over its spin axis, a process that is increasingly accepted as a major tectonic force in shaping crustal configurations [2], and as suggested here, in biological history as well. The ammonite fossil record from James Ross Island shows an increase in species from the Santonian and Campanian Stage strata there, an increase that is in part composed of the appearance of warmer water species of the Indopacific Biotic Realm, coupled with an apparent increase in paleotemperature of the basin over the same time interval. These separate lines of evidence are consistent with our hypothesis of a rapid northward movement of Antarctica during the Santonian Age, taking the James Ross Basin habitat into warmer waters, and into marine ecosystems that also were inhabited by Indopacific Province ammonites. Their equally rapid extinction several million years later is also consistent with the mirror image return of Antarctica (TWP movements are composed of a couplet of movements in opposite directions) back into higher latiutudes during the Campanian Age, and in so doing into colder water, with cooling presumably the cause of the rapid extinction of the Indopacific fauna. The later re-population of ammonites from the well known beds on Seymour Island we also attribute to a slow, plate tectonic-driven movement into lower latitude during the Maastrichtian Age, as evidenced by the combined, Coniacian throuth Maastrichtian paleomagnetic record that we document here for the first time.
Subject Terms
Geophysics, Paleobiology, Paleoclimatology
Introduction
The thick and well-exposed, Upper Cretaceous beds cropping out in the James Ross Island region, Antarctica (including James Ross, Seymour, Vega, and Snow Hill Islands, Figure 1) may be the most expanded, fossiliferous beds with both micro- and macrofossils of Late Cretaceous-Early Paleogene age yet know anywhere in the globe. They also represent the southermost known KPg boundary on land. As such, these Antarctic beds provide unparalleled opportunity to test hypotheses concerning the pattern, cause, and consequences of the KPg mass extinction in the Southern Hemisphere, including events and fossil ranges leading up to the terminal Cretaceous event, the event itself [3-5], and its aftermath, as well as documenting biotic events that occurred in zones and stages prior to, and after the K/Pg global mass extinction.
The fossil record for the ~20 million years prior to the KPg mass extinction event has yielded information about diversity and biogeography that remains both interesting and relatively unexplained [6-9]. These are the reason(s) for observed trends in ammonite diversity, disparity, and biogeographic affinities in fossils from the Santonian and Campanian Stages in the MG basin, as well as reasons for the rapid extinction of this fauna. However, to date there has been no means of high resolution chronostratigraphy allowing identification of the Coniacian, Santonian, and Campanian stage and substage boundaries, and this lack of temporal precision has limited the resolution of correlation to lower latitude, possibly global events that pre-dated the KPg mass extinction.
Here we use the first Coniacian through Campanian-aged magnetostratigraphy from Antarctica to identify stage and substate boundaries, and then use these to better constrain the age of the curious ammonite record obtained from James Ross Island. That record demonstrates that there was a rapid ammonite diversification beginning in the Santonian age, anomalous in its biogeographic affinity, and that this Indopacific fauna was entirely extinct by the end of the early Campanian age, or by about 80-78 mya [3-5].
As shown in Figure 2, representatives of scaphitid, baculitid, and nostoeratid amonitesas well as inoceramid bivalves whose families all disappeared at the KPg boundary in the northern hemisphere disappeard in non-sychronous fashion in the JRI basin as much as ten million years before the KPg boundary event [6,8,10,11]. As yet there remains no viable hypothesis explaining these early extinctions. Equally problematical is the rapid diversification of ammontes of Indopacific Province affinity during the Santonian Age.
Materials and Methods
We sampled for magnetostratigraphy and biostratigraphy on James Ross Island in the (from oldest to youngest) Hidden Lake, Santa Marta, and Snow Hill Island Formations; Figure 1. We located and used previously published, measured stratigraphic sections [6-9] for our sampling that were kindly pointed out to us in the field by Dr. E. Olivero, so as to be able to integrate his previous fossil collection and biostratigraphy with our magnetostratigraphic and ammonite specimen sampling. The longest continuous outcrop extends from the shores of Whiskey Bay up to the summit of Santa Marta Pass, in the Santa Marta Foramation. We also sampled older strata of Hidden Lake Formation as shown in Figure 1. From the top of Santa Marta pass, we further sampled Santa Marta beds in two, non contiguous sections, one starting from near the pass, one on the shore of Brandy Bay itself.
From oldest to youngest, these sections included H4, M0, G120, G8, F16, D8, and 14S. At each site we located the base of the section as described in previous work [6-9] and then re-measured each section using Jacobs staff fitted with Abney Level. All sample levels come from our re-measures. Strike and dip was taken at each locality. The majority of samples were drilled into concretions, in a small number, where drilling was not possible, we used plastic box samples covering oriented samples carved from a single bed. Each site that was sampled (magnetic cores or box samples taken) had GPS information recorded for it, and this information available upon request.
The sampled sections produce a nearly continuous outcrop that atextends from the shores of Whiskey Bay up to the summit of Santa Marta Pass, thus extending from the Hidden Lake Formation through much of the Santa Marta Formation. However, strata on these transects were often covered by overburden. In general there was more exposure on the Whiskey Bay side of Santa Marta Pass (Sections H4 to near the top of F16), but less coverage from F16 through D8; A pronounced missing interval separates these two. Also, the sections of HL and MO were isolated, and their relative stratigraphic positions one to another (relative to their base and tops) was not clear in the field.
For magnetostratigraphic sampling, we took either block samples (a minority) or more frequently used a specially built gasoline powered drill (modified from a chain saw) that took 2.5 cm wide cores up 5-8 cm deep into the rock. Most cores were taken from concretions, which were common throughtout both formations sampled. These cores were oriented using both a sun compass (when sun was available) and a magnetic compass, then extracted from the rock. Samples were taken to the paleomagnetics lab at the California Institute of Technology (Caltech), part of the RAPID consortium of paleomagnetics labs.
In the laboratory, cores were cut into slices, and slices furthest from the weathering surface were analyzed preferentially. Natural remanent magnetization (NRM) was measured in both the up and down positions using automated techniques of the Rock and Paleomagnetic Instrument Development consortium (RAPID) system. Samples were then subjected to several sequences of alternating immersion in liquid nitrogen and remenance measurement to remove viscous components held my multi-domain magnetite. Subsequently, they were subjected to increasingly strong low alternating field demagnetization up to roughly 7.0 mT to remove spurious signals that may be introduced by airport X-ray machines and the like. Finally, samples were then treated with thermal demagnetization in an oxygen-free (pure N2) atmosphere, alternating oven heating and remanence measurements from 60 ℃ until they started displaying unstable behavior between ~250 to 500 ℃. From our earlier paleomagnetic work on Seymour Island, [1] we found that most secondary components were removed below these levels. Examples of sample behavior of palemagnetic cores from the basin have previously been illustrated [1].
These results were then quantified by standard principle component anlaysis [12]. Previous work using a fold test on Seymour Island [1] indicated that results obtained were primary, Cretaceous-aged directions, rather than modern field overprint. This previous work used secondary samples of all litholgies to determine magnetic mineralogy, including anisotropy, IRM, ARM, RRM, and backfield measurements, using standard laboratory methods [13].
The Inclination results from our interpretable samples can be placed in their stratigraphic order to provide a magnetostratigraphic record for our sampled sections. When combined with biostratigraphic information, the magnetostratigraphy provides the highest resolution chronostratigraphic framework possible in the JRB to date, and allows completion of the Santonian through Maastrichtian Age magnetostratigraphic record in the James Ross Basin.
Results
Magnetostratigraphy
The inclinations and declinations of our samples are listed in Table 1. The positions of the chron reversals allowed us to correlate to the Global Polarity Time Scale, and thus to stage boundaries and substage boundaries. The positions of our measured sections are shown in Figure 1, and their correlation to the GPTS based on our magnetostratigraphic results is shown in Figure 3, Figure 4, Figure 5 and Figure 6.
The lowest reversal was observed in section HL, although there are also normal directions from this section, suggesting that these early reversal results from HL might be spurious. The ammonite record of lower Santonian baculitids suggests that this section, as well as section MO, and reversed polarity was continuous through sections G120, the superjacent G8, and through most of superjacent F16. The geographic separation of HL from H4 and M0 has Normal polarity was observed high in section F16, and then was continuous through section D8 (although magnetic behavior of many of the cores from the latter precluded accurate polarity determination. Both well behaved normal and reversed polarities were observed in section 14S.
The sequence of Chrons extends from an indeterminate level within the Cretaceous "Long Normal" (Chron 34N) up to the vicinity of the Campanian-Maastrichtian stage boundary as understood in the European type regions, which corresponds to the top of the Nostoceras hyatti ammonite zone. The lowest reversal boundary (interpreted here as the 34N/33R boundary) recognized here was in the vicinity of the Hidden Lake - Santa Marta Formation boundary, while the superjacent, 33R/33N boundary was found near the Santa Marta Pass, but within the Santa Marta Formation. The highest reversal boundaries, 33N/32R, and 32R/33N, were found in the Snow Hill Island Formation, in section 14S; these positions related to lithostratigraphy and biostratigraphy are shown in Figure 4 and Figure 5.
Reversal boundaries and biostratigraphy
Our oldest samples (Hidden Lake Formation, from section HL) are of normal polarity, and, as noted above, are overlain by a thick succession of reversed polarity, making up most or all of the Santa Marta Formation on James Ross Island. These lowest samples (in section HL) we interpret as correlative with magnetochron 34N, the highest part of the "Cretaceous Long Normal", because of the presence of Santonian-aged fossils (among ammonites [6] and inoceramid bivalves [14], among other taxa, in the high part of the Hidden Lake Formation and the basal Santa Marta Formation. Samples from H4 and MOT showed both normal and reversed polarity, suggestive of the base of Chron 33R in the highest parts of both sections.
The strata of normal polarity are overlain by a thick interval of reversed polarity of the Santa Marta Formation; the greatest part of this formation, in fact, is reversed being composed of strata deposited during Chron 33R. Near the top of that formation, near the top of section F16, we recovered normal polarity once again. We interpret this as the 33R-33N-chron transition, which would make it correlative to the recently described transition from Reverse to Normal Polarity from another, but isolated section on James Ross Island [15], which has yielded an ammonite fauna that is equivalent to the Baculites inornatus assemblage recognized from our study area, as well as to the ammonite faunas of Sucia Island, Washington, and the slightly younger fauna from Punta San Jose, Baja California, both with published magnetostratigraphy [16,17].
Biostratigraphic support for interpreting the thick sequence of reversed polarity through sections G120, through G8, and through most of F16 as Chron 33R is based on the presence of cosmopolitan ammonite species first identified as having strong affinities with biochronologically important species of the Indopacific Province [6-8]. Among these are Baculites bailyi, B. rectus (which here we identifiy as B. chicoense), and B. inornatus. All of these are found in sections with reversed polarity in the Indo-Pacific Province sections of California and British Columbia [16].
Baculites inornatus is found both in chron 33R in California and Washington State, as well as in the succeeding chron 33N in Baja California [16,17]. Baculites subanceps, also identified both in the Larson Basin and the Pacific Coast of North America and Japan, is found in both 33R and 33N in North American deposits, as shown in Figure 6.
Our results largely confirm previous results [7] concerning the disappearance of baculitid ammonites as occurring either high in chron 33R or more probably low in 33N; the final disappearance of inoceramids in 33N (although there is a prior extinction of sphenoceramids and platyceramids (Inoceramidae) early magnetochron 33R), and disappearance of scaphitid and nostoceratid ammonites in chron 33N [7-9].
Finally, the discoveries [7] of both Hoplitoplacenticeras sp. and Metaplacenticeras sp. near the top of section F16 [7], is strong evidence that the top of the Santa Marta Formation (top of F16) is being correlative with the approximate time interval represented by the chron 33R-33N transition. These also tie the these Antarctic sections to both the Nanaimo Group of British Columbia and the Great Valley Sequence of California) [16,17], as well as to the newly reported results from Hamiliton Norte and Redondo Point, James Ross Island [15].
While sample behavior worsened near the top of section F16, and was poor for about half the cores from D8, there is enough information to constrain the chron boundary to the top 20-30 m of section F16.
From the base of section D8 upward, there is increasing cover, and little fossil record in the stratal layers that are exposed. Between (or perhaps partially overlapping) section D8, and the base of section 14S there is a further fossiliferous section, section QF, that unfortunately appears to be a slump block and not in place. It was not sampled for magnetostratigraphy. However, section 14S, exposed along the shoreline of Whiskey Bay, was sampled, and is extremely important for both chronostratigraphic as well as paleoecological results. This approximately 60 m thick section was exceptionally fossiliferous, and is also the sole place in our study on James Ross Island where unaltered aragonitic fossils could be recovered. We found a final, normal-to-reversed polarity change in the section 14S. However, at least 200 m of covered section separates the highest samples of section D8 from the lowest of 14S.
This section (14S) represent the highest Campanian beds on James Ross Island. This section is represented graphically as continuous with the long section of Santa Marta Formation, and lowest Snow Hill Island Formation beneath it [7]. Based on the magnetostratigraphic results, it is older (lower) than the Campanian-Maastrichtian boundary, and probably one zone older than even the last ammonite zone of the Upper Campanian in the Northern Hemisphere, the Nostoceras hyatti Zone. This locality is entirely within the Snow Hill Island Formation, and is of both reversed and normal polarity, placing it within the C32R-32N chron boundary transition. While ammonites in section are numerous, (and large; virtually all are planispiral ammonites up to 1.3 m in diameter), their diversity is low: only two species, and all other invertebrate fossils are rare. The most common species, earlier identified as Anapachydiscus sp. 3) [6] is, based on our interpretation, a homeomorph (or is) the well-known Indopacific species Anapachydiscus peninsularis.
In Baja California, at the locality known as San Antonio del Mar, this species ranges through part of Chron 32R [16]. Thus for structural (position within the succession) and biostratigraphic reasons. this reversed interval on James Ross Island is most parsimoniously identified as 32R, making it the same age as the well-studied beds of Hornby Island, Vancouver Island region, that contain Baculites occidentalis and Nostoceras hornbyense as well as in the Rosario Formation of Baja California.
Ammonite diverisity and identifications
The identities of the ammointes observed in this study are the result of the superb, lifetime work of Eduardo Olivero, who not only collected the vast majority of specimens from James Ross island, but was the first to recogonize the appearance of Indopacific species, and the species shown and their ranges are mainly the product of Olivero and his co-workers. The only significant change we propose is that Baculites rectus is a junior synonym of B. chicoense. This change, however, is important, and allows a further erection of a zonation based on cosmopolitan, Indopacific ammonite species that can augment the current biostratigraphy, that is based mainly on endemic species. The only morphological difference between Baculites rectus and B. chicoense is that most of the James Ross Island specimens lack the ventral keel that many (but not all) of B. chicoense from California and the Nanaimo Group possess. Howerver, recent work [16] showed that presence or absence of a keel is not sufficent to serve as a single character that idnetifies a specific species. Keels were observed to appear in changing percentages of both B. inornatus from the Nanaimo Group, California, and the Rosario Formation of Baja California, as well as among specimens of B. gregoryensis of the Western Interior. The recognitiion of B. chicoense from James Ross Island is not particularly surprising because of the presence of many other species of western, North America affinity amid the strata of Chron 33R on JRI, as shown in Figure 6, based on prior identifications [6-8] and our own identifications of ammonites. The complete diversity of ammonites we recognize from JRI, tied to the GPTS is shown in Figure 6.
Inclination, diversity and paleotemperatures
We show the relationship bewtween ammonite diversity, inclination, and previously published paleotemperatures in Figure 7. These data show that the rapid diversification of ammonites coincided with a reduction in paleolatitude (toward warmer water), while the reverse of this trend, starting in the Lower Campanian, coincided with the observed Campanian extinctions. The new chronostratigraphic data also confirms that the time interval of these ammonite diversity changes was consistent with the time interval previously identified for a Late Santonian into early Campanian TWP event. The rate of movement of a 20° Santonian TPW event that was identified from other regions [18] indicates no less than a half meter per year movement, compared to the plate tectonic speed limit of less than 20 mm/year.
Our data indicate that maximum ammonite diversity corresponded to both the most outward (lowest latitude) position as well as the highest measured temperatures in the MG basin [19], while subsequent diversity lows from the Upper Campanian to the Upper Maastrichtian all took place while the MG was at or above 70° South. The significant relationship between MG paleolatitude and ammonite diversity that our data has revealed is shown in Figure 7.
Finally, there was a new movement toward lower (warmer) latitudes in the Late Maastrichtian, one that also corresponded to both an ammonite diversification, as well as temperature increase in the basin (Figure 7). However, this Late Maastrichtian ammonite fauna never reaches the number of species from the basal Campanian, maximum outboard position, and in comparison to other regions with latest Maastrichtian ammonites is of lower diversity. These species, including the important, zonal index pachydiscid as well as kosmatticeratid ammonites are all endemics.
The same trend may have occurred in inoceramid bivalves. In the Upper Santonian beds on James Ross Island, we collected numerous specimens of mytiloid and platycermid bivalves belonging to the Inoceramidae. Both are morphologically similar to the Indopacific species Inoceramus (Mytiloides) naumanni and I.(M). orientalis, as well as Inoceramus (Platyceramus) ezoensis, indicating that like the ammonites, an Indopacific inoceramid fauna was present. However by the early Campanian ammonite diversity crash, the Indopacific inoceramid fauna also went extinct, and probably for the same reason. Only in the Upper Campanian is there a new record of an inoceramid in the basin, but it was short lived, endemic, and highly unusual in morphology compared to the Santonian, Indopacific inoceramids. For all practical purposes, the extinction of the (typical) inoceramids occurred early in Chron 33R.
Among the early extinctions, from strata now magnetostratigraphically dated, were scaphitid, baculitid, and nostoceratid ammonites (including species of the Indopacific (or global) genera Baculites, Polypychoceras, Eubostrychoceras, Ryugasella, Ainoceras,Pseudoxybeloceras, and at least two different species of coleoids. These "early" disappearances were unique for the Late Cretaceous marine world as has been previously pointed out by Antarctic paleontologists as referenced above. To date there have been no explanatory hypothesis, other than generalized suppositions that temperature change,or oxygenation of the deeper basinal waters, may have been been involved. None of these seem viable [20].
Re-diversification of ammonites did not occur until the start of the Late Maastrichtian, and even then the diversity and more importantly the assemblage of ammonite morphotypes of this last Antarctic ammonite fauna was different from all other known Late Maastrichtian ammonite faunas, which elsewhere have significant numbers of scaphitid, diplomoceratid, nostoceratid, and baculitid heteromorphs. For example, in the Bay of Biscay, the best studied Late Maastrichtian to K-Pg boundary site known, over 25% of the fauna (33 species) were heteromorphic, compared to one out of the only nine species of Late Maastrichtian ammonites from Antarctica.
Discussion
Because of the presence of Indopacific ammonites in Antarctica, an important question is whether the ranges in both places are similar, and if not, which occurred first: Did the species first appear in Antarctica and migrate north, or vice versa? Vertebrate paleonotologists are asking similar questions: Was the JR Basin a cradle of origination?
The ranges of Nanaimo Group ammonites over the approximate Santonian through Campanian from the Vancouver Island (Figure 6) region has the best stratigraphic range information of anywhere in the Indopacific Province. The recent discovery that Chron 33N extends down into the Elongatum Zone of both the Vancouver Island as well as Sacramento Valley (Forbes Formation and Chico Formation; Mitchell, et al. in Review) [21] appears to be significantly different than its lower range on James Ross Island, where the index species is entirely contained in 33R, indicating that its range in the Indopacific Province might predate its first appearance in the Antarctic province. The majority of the Elongatum Zone in North America is within Chron 34N, whereas it does not appear in normal polarity beds on JRI, indicating that it arrived only later in Antarctica.
The only other difference (if indeed there is one with Eubostrychoceras elongatum) in ranges based on presence or absence of chrons could be for Baculites inornatus, which is known to range across the Chron 33R -33N transition, appearing in the Rosario Formation in normal polarity, but in reversed polarity elsewhere in the province (Sucia Island, of the Nanaimo Group, and Sand Creek and elsewhere in the Forbes Formation of the Sacramento Valley [16,17]. On JRI, all specimens of B. inornatus come from reversed polarity (Chron 33R). However, the large covered interval between sections D8 and 14S could very well obscure a similar higher range of this taxon in the Antarctic Province.
To date there has been no single (or multiple hypotheses) that could explain the biogeographic, diveristy, and extinciton patterns of ammonites in the Marambio Basin during the Santonian through Maastricthtian Age time interval that seem viable. There are only guesses. One such that we propose here is that the circum- Antarctic current was present in the late Cretaceous (Santonian, shut off for the late Santonian into Campanian) and then started again, with its presence essential fencing off the MG basin from any other kind of faunal exchange. However, there is abundant geological evidence that this circum-Antarctic current did not start up until age estimates of 32 to 35.5 Ma [20], and in any case this cannot explain the paleolatitudinal data reported here. That there was a TPW event seems more likely.
Until now, two large scale biotic process (pre-KPg extinctions, and a short-lived interval of changing biogeographic affinity in the basin), and one physical change (a major paleotemperature change independently reported affecting the Marambio Group (Late Cretaceous-Eocene) of the Antarctic Peninsula have been treated as independent, and both have remained unexplained for decades, beyond unsatisfactory generalization about changing sea level and ocean temperatures caused by gradual climate change over this time interval. The data presented here provide evidence for a paleobiologically significant TPW event, and in so doing finally answer long running paleobiological questions about the Late Cretaceous macrofauna of Antarctica. There is previous evidence that such an event occurred in the Late Cretaceous [18], and prior paleotemperature analyses indicate warmer temperatures during the late Santonian than from the Maastrichtian fossils studied to date [19].
The James Ross Island data also can now be appended to the prior, published magnetostratigraphic record of the Maastrichtian on Seymour Island to present a combined, Santonian through Maastrichtian paleomagnetics record for the James Ross Basin Upper Cretaceous. While there remain "holes" in this record, most noticeably the exact placement of the top of chron 33R, in fact the all post 34N, "long normal" chrons of the Upper Cretaceous can now be recognized, which allows a new, independent dating scheme for testing hypotheses of global biotic and environmental change in high southern latitudes for the end of the Cretaceous.
It is the latter series of bioevents that are perhaps the most interesting development to come from the biostratigraphic studies of Late Cretaceous fossils in the James Ross Island Basin. These include what appear to be both anomalously early appearances of groups (such as the early appearance of the otherwise, Late Maastrichtian planktonic foraminifera Abathomphalus mayaroensis from Campanian strata) as well as the early extinctions of fossil groups that elsewhere ranged higher in time. For instance, the disappearance of inoceramid bivalves occurred earlier in Antarctica than anywhere else in the world, as did the disappearance of both baculitid and nostoceratid ammonites. By the Nostoceras hyatti zone, the highest zone of the Campanian with almost worldwide extent and a zone that in time immediately succeeds our highest magnetostratigraphic samples on James Ross Island, the ammonite faunas of the Antarctica region appear to have been totally cut off from the rest of the world, fueling speciation of endemics. This changeover to a highly endemic assemblage of zonal index fossils, and because of the lack of aragonite from James Ross Island Late Cretaceous aged strata has precluded the use of Sr isotope dating, and is in large part a reason that independent age control for verifying age calls at even a substage level have been important.
While the Santonian-aged species noted from the lowest Santa Marta beds have been given names identified from southern, Indopacific-aged inoceramid taxa, we find (although none of us are specialists on inoceramids) that the Santonian-aged inoceramids appear morphologically indistinguishable from Sphenoceramus orientalis, and Platyceramus ezoensis, (Figure 4) both of which are found in California and the Vancouver Island region with Eubostrychoceras elongatum, Eupachydiscus sp. aff. E. haradai, and Baculites baileyi in both North Pacific, Indo- Pacific, and Antarctic strata. Our new magnetostratigraphic data thus provides and opportunity to test whether these co-occurring species have similar time ranges or are diachronous in FADs or LADs. It appears, in fact, that the appearance and disappearance of these taxa relative to the Chron 33R boundary are synchronous in both places.
The most striking aspect of the ammonite record from the Santonian-through Campanian strata of James Ross Island concerns the rapidity of diversity in the high Santonian, and and equally rapid (and equally puzzling diversity drop between the rich and diverse ammonite fauna below our section D8, and from D8 all the way up to the Upper Maastrichtian of Seymour Island [22].
For the first time, the presence of a shared magnetostratigraphic record allows comparison of the JRI diversity drop to coeval time in the Indopacific Basin. Using Figure 6, it appears that there is also a diversity drop (albeit of less magnitude in the Nanaimo and Sacramento Basins. The rich and diverse fossils of the Vancouverense Zone in both places is followed by the extremely low diversity assemblages of Chron 33N in both Nanaimo and Sacramento Valley, on both sides. For instance, the fossil rich assemblage of low 33N at Punta San Jose contains no planispiral ammonites at all [16,17]. Similar strata on Vancouver Island also shows a marked diversity drop late in Chron 33R and into 33N. These beds are overlain by a low diversity assemblage with mainly Metaplacenticeras spp. for some millions of years, prior to the advent of the return to high diversity present in California (the Pachydiscus Silt of the San Joaquin Valley) and Hornby Island, Nanaimo Group. Whether this is an effect of sea level, or other, perhaps global events remain to be seen.
What remains enigmatic are 1) The anomalously high diversification of ammonites, 2) Of a different biogeographic realm than was present in Coniacian time, that 3) Occurred in a short interval of geological time and, was 4) Accompanied by evidence of rapid warming, followed by 5) Equally rapid extinction, and 6) Cooling. The movement of Antarctica during the Santonian and early Campanian, as shown in Figure 8, is a viable hypotheiss explaining all of these lines of evidence.
Acknowledgments
We would like to thank our funders, Polar Programs, National Science Foundation (PLR 0739432, Co-PIs Kirschvink, Steig, Ward), and National Science Foundation Geophysics, # EAR 1114011PI Ward. Great help was given by the officers and crew of the RV Lawrence Gould. This work could not have been completed without the help of Dr. Eduardo Olivero and the Argentinian Antarctic program. Thanks to Tom Tobin for running the magnetic core samples while at Cal Tech, to Joe Kirschvink, Tobin, Shane Schoepfer, Kelly Hillbun, and David Smith for help in the field. Tom Tobin drafted Figure 1.